
人衰老成纖維細(xì)胞經(jīng)紫外線損傷后的DNA 修復(fù)和細(xì)胞周期調(diào)控
人衰老成纖維細(xì)胞經(jīng)紫外線損傷后的DNA 修復(fù)和細(xì)胞周期調(diào)控
段建明 張宗玉 童坦君
(北京醫(yī)科大學(xué)生物化學(xué)與分子生物學(xué)系, 北京 100083)
摘要 以體外培養(yǎng)的不同代齡的人胚肺二倍體成纖維細(xì)胞(2BS) 為對象, 紫外線誘導(dǎo)DNA 損傷后, 觀察細(xì)胞形態(tài)、增殖特性、細(xì)胞周期、DNA修復(fù)變化等細(xì)胞應(yīng)答以及gadd153、p21、p53 等基因的轉(zhuǎn)錄水平的表達(dá)變化. 結(jié)果顯示: 紫外線誘導(dǎo)DNA損傷后, 衰老(>55代) 2BS 細(xì)胞形
態(tài)及增殖能力的改變不如年輕細(xì)胞(<30代) 顯著; 不同代齡的細(xì)胞損傷后均出現(xiàn)G1 期阻滯現(xiàn)象, 年輕細(xì)胞G1 期阻滯率明顯高于衰老細(xì)胞(P<0105) ; 衰老細(xì)胞總的修復(fù)能力較年輕細(xì)胞明顯下降(P<0101) ; 同時(shí), gadd153、p21、p53 等的可誘導(dǎo)性均低于年輕2BS 細(xì)胞. 由此, 分別在細(xì)胞水平與基因水平反映了衰老細(xì)胞經(jīng)紫外線照射損傷后的細(xì)胞應(yīng)答變化與修復(fù)機(jī)能減退的關(guān)系.
關(guān)鍵詞 衰老細(xì)胞,DNA 損傷修復(fù), 細(xì)胞周期, 檢驗(yàn)點(diǎn)控制
DNA 損傷的積累及修復(fù)能力的下降是生物衰老的重要原因之一[1]. p53、p21 和ATM 是近年發(fā)現(xiàn)的人類細(xì)胞3個(gè)DNA 損傷應(yīng)答的檢驗(yàn)點(diǎn)控制(check2point control) 基因[2]. p53、p21 不僅作為抑癌基因發(fā)揮重要作用, 且在DNA 損傷的細(xì)胞應(yīng)答中起中心調(diào)控作用, 它們可調(diào)節(jié)DNA損傷修復(fù)、細(xì)胞周期阻斷以及細(xì)胞凋亡等諸多基因的轉(zhuǎn)錄[3]. DNA損傷可誘導(dǎo)基因中的gadd45、gadd153與DNA 損傷后的細(xì)胞周期阻滯以及修復(fù)也密切相關(guān), 可能作為p53的下游基因而起作用[3]. 這些基因在衰老細(xì)胞中有無表達(dá)變化及與衰老細(xì)胞修復(fù)能力的下降是否有關(guān)? 至今未見國內(nèi)外報(bào)道. 我們以體外培養(yǎng)的人胚肺二倍體成纖維細(xì)胞(2BS)為對象, 研究了紫外線照射后衰老細(xì)胞中的DNA 損傷可誘導(dǎo)基因gadd153 以及檢驗(yàn)點(diǎn)控制基因p21、p53的表達(dá)變化,及其與DNA修復(fù)能力和細(xì)胞周期變化的相關(guān)性.
1 材料與方法
1.1 材料及主要試劑 人胚肺二倍體成纖維細(xì)胞引自衛(wèi)生部生物制品研究所, pB luesSK 質(zhì)粒(gadd153 cDNA ) 由北京醫(yī)科大學(xué)人民醫(yī)院惠贈,p21WAF1.CIP1. SDI1cDNA由美國Baylor 醫(yī)學(xué)院惠贈,p53及B2act in cDNA 探針為本室留存. DMEM干粉培養(yǎng)基: GIBCO.BRL公司, 胎牛血清: 北京北郊農(nóng)場血液制品所, 3H2TdR 及A232P dCTP: 北京亞輝生物工程公司, P rim e2a2Gene Lebeling System:Promege公司, 羥基脲、鮭魚精DNA、聚乙烯吡咯烷酮、牛血清白蛋白: Sigma公司, 其它試劑均為Sig2ma產(chǎn)品或國產(chǎn)分析純.
1.2 細(xì)胞培養(yǎng) 人胚肺二倍體成纖維細(xì)胞(2BS)培養(yǎng)于含10% 胎牛血清的DMEM 培養(yǎng)液中, 37℃溫育箱中生長, 傳代至62±4 代及24±6 代分別作為衰老及年輕細(xì)胞使用.
1.3 細(xì)胞形態(tài)及增殖特性觀察 細(xì)胞生長至對數(shù)生長期后, 細(xì)胞懸液調(diào)整至1×104細(xì)胞.ml, 移入24孔板中培養(yǎng), 待細(xì)胞貼壁后以紫外線(0145J.m 2.s) 照射5 min 損傷細(xì)胞[4],更換新鮮培養(yǎng)液于37℃溫育, 每12h觀察細(xì)胞形態(tài)變化并進(jìn)行細(xì)胞計(jì)數(shù), 以未進(jìn)行紫外線照射的衰老及年輕細(xì)胞作對照.
1.4 細(xì)胞周期分析 紫外線照射劑量、時(shí)間同上,37℃溫育24h后胰酶消化, 收集細(xì)胞, 再用RNA 酶于37℃消化1h, 然后用碘化丙啶(PI)染色, 以Bec2ton2Dickinson 流式細(xì)胞儀進(jìn)行FACScan(fluo res2cence act ivated cell sorting)分析, 以未進(jìn)行紫外線照射的衰老及年輕細(xì)胞作對照。
1.5 UDS (un scheduled DNA syn thes is) 測定 細(xì)胞計(jì)數(shù)后, 以1×105 細(xì)胞.孔傳于6孔板, 待細(xì)胞完全貼壁后,以含015% 胎牛血清的DMEM培養(yǎng)液饑餓細(xì)胞72h使細(xì)胞同步化,更換含5mmo l.L羥基脲的無血清DMEM 培養(yǎng)液于37℃溫育1h后,紫外線照射, 其劑量與時(shí)間同上,更換培養(yǎng)液(含5mmo l.L 羥基脲、1LCi.m l3H2TdR、10%胎牛血清的DMEM) , 37℃溫育不同時(shí)間收集細(xì)胞,進(jìn)行3H2TdR計(jì)數(shù), 對照組為未進(jìn)行紫外線照射的細(xì)胞[5]。
1.6 斑點(diǎn)及Northern 印跡雜交細(xì)胞處于對數(shù)生長期時(shí)進(jìn)行紫外線照射,時(shí)間、劑量同上, 然后于37℃溫育不同時(shí)間收集細(xì)胞, 用異硫氰酸胍一步法提取細(xì)胞總RNA,然后轉(zhuǎn)膜、標(biāo)記探針及進(jìn)行斑點(diǎn)和Northern 雜交[6,7] , 以未進(jìn)行紫外線照射的細(xì)胞為對照. 雜交結(jié)果由積分光密度掃描分析。
2 結(jié)果
2.1 細(xì)胞形態(tài)及增殖特性的比較 紫外線照射前可見衰老2BS細(xì)胞形態(tài)與年輕細(xì)胞明顯不同, 衰老細(xì)胞胞體肥大、胞漿內(nèi)顆粒增多、胞漿混濁、折光性下降、細(xì)胞排列極不規(guī)則; 而年輕細(xì)胞則胞體細(xì)長、胞漿清亮、細(xì)胞呈漩渦狀排列. 紫外線照射后, 衰老細(xì)胞形態(tài)變化不十分明顯, 僅見胞漿內(nèi)顆粒略顯增加; 而年輕細(xì)胞則可見胞漿顆粒明顯增多、胞漿透明度下降、胞體也略顯增大, 但培養(yǎng)約1周后細(xì)胞形態(tài)基本恢復(fù)正常. 紫外線照射前后細(xì)胞生長曲線見Fig.1, 經(jīng)紫外線照射后年輕細(xì)胞增殖速率受到明顯抑制, 但約6d后增殖能力恢復(fù)正常; 衰老細(xì)胞增殖速率明顯低于年輕細(xì)胞, 紫外線照射前后增殖變化不十分明顯。
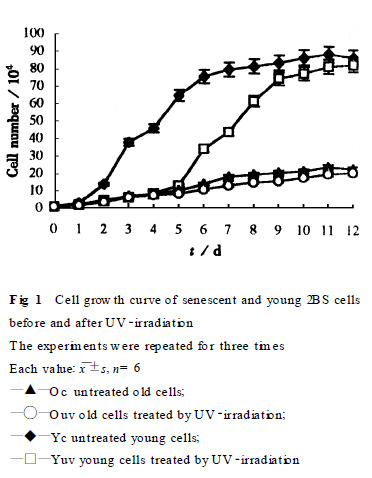
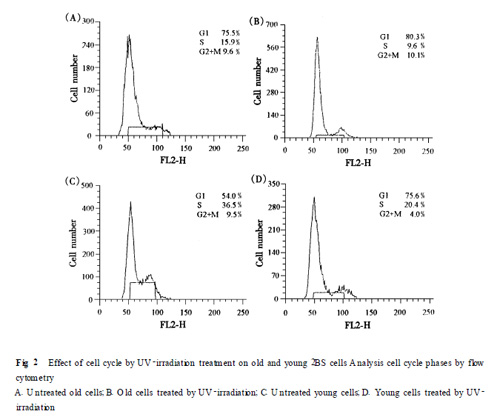
212 細(xì)胞周期時(shí)相變化 紫外線照射后衰老及年輕的2BS 細(xì)胞均可見G1 期阻滯, 而年輕細(xì)胞G1 期細(xì)胞阻滯率高于較衰老細(xì)胞(P<0105) (Fig.2).
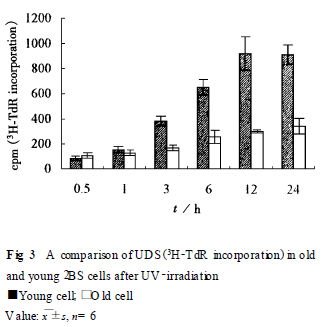
213 DNA 修復(fù)能力比較 以3H2TdR 參入法測定非程序性DNA 合成(UDS) , 以表示DNA 總修復(fù)能力. 從F ig. 3 可見, 在紫外線照射后12 h 左右UDS達(dá)峰值. 衰老2BS 細(xì)胞的修復(fù)能力明顯低于年輕細(xì)胞(P < 0101).
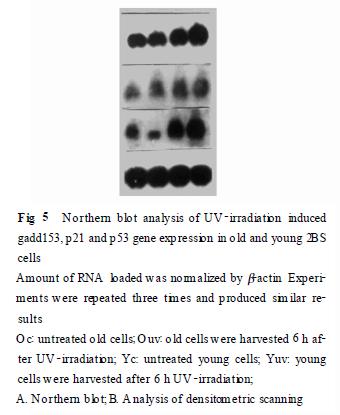
214 斑點(diǎn)及Northern 印跡雜交結(jié)果分析 由斑點(diǎn)雜交結(jié)果可見, 紫外線照射6 h 后gadd 153 表達(dá)達(dá)峰值, 衰老細(xì)胞的gadd 153 可誘導(dǎo)性明顯低于年輕細(xì)胞, 結(jié)果見(Fig. 4). 根據(jù)斑點(diǎn)雜交結(jié)果選取紫外線照射后6 h 收集細(xì)胞, 進(jìn)行Northern 雜交, 結(jié)果顯示(Fig. 5) , 紫外線照射后衰老細(xì)胞中g(shù)add153、p 53、p 21 基因的可誘導(dǎo)性均明顯低于年輕細(xì)胞, 但其中p 53 誘導(dǎo)性改變最小. 另外, 這三種基因在紫外線損傷前的基本表達(dá)在衰老細(xì)胞及年輕細(xì)胞中也有所不同, 其中p 21 在衰老細(xì)胞中的基本表達(dá)明顯高于年輕細(xì)胞, gadd153 的表達(dá)在衰老細(xì)胞中似略高于年輕細(xì)胞, 而p53 的表達(dá)差異不顯著.

3 討論
細(xì)胞衰老是一個(gè)多因素、多途徑所致的復(fù)雜結(jié)果,DNA 損傷的積累及修復(fù)能力的下降可能是細(xì)胞衰老的重要原因之一[8] , 在DNA 受損后衰老細(xì)胞應(yīng)答與年輕細(xì)胞有所不同. 衰老細(xì)胞的形態(tài)及增殖曲線的改變不明顯, 同時(shí)其恢復(fù)也緩慢. 流式細(xì)胞計(jì)檢測結(jié)果顯示, 衰老細(xì)胞G1 期阻滯率的增加幅度低于年輕細(xì)胞; 同時(shí)衰老細(xì)胞的DNA 修復(fù)能力顯著低于年輕細(xì)胞. 這些結(jié)果都說明衰老細(xì)胞對于DNA 損傷的應(yīng)答能力明顯不如年輕細(xì)胞. 那么, 衰老細(xì)胞對DNA 損傷的應(yīng)答能力下降的根本原因是什么呢? p53、p21 和gaddl53 等基因表達(dá)狀況在紫外線照射前后的變化可能為我們提供了一些有價(jià)值的線索.
DNA 損傷后, 會通過一定的信號傳導(dǎo)途徑引起細(xì)胞反應(yīng), 誘導(dǎo)眾多基因表達(dá)的激活或抑制,DNA 修復(fù)系統(tǒng)的激活則是細(xì)胞維持自身功能完整性和細(xì)胞生存的一個(gè)重要機(jī)制[ 9 ]. p 53 在DNA損傷監(jiān)控中處于檢驗(yàn)點(diǎn)控制的位置, 起著中樞作用,p21 及gadd153 均為其下游基因, 它們有著部分共同的途徑, 又可通過各自不同的途徑作用于細(xì)胞周期及修復(fù)系統(tǒng), 在DNA 損傷修復(fù)基因調(diào)控網(wǎng)絡(luò)中既互相協(xié)同又相互影響, 發(fā)揮著各自不同的作用[9]. p21是周期蛋白激酶的抑制因子,作為p 53的下游基因, 在p 53與細(xì)胞周期調(diào)控之間起橋梁作用. p 21的高表達(dá)會導(dǎo)致細(xì)胞周期阻滯, 還會通過與PCNA (增殖細(xì)胞核抗原) 相作用影響DNA 的復(fù)制及其修復(fù)[3]. gadd153 在紫外線的誘導(dǎo)表達(dá)中也是依賴于p53的, 它作為p53 的下游基因,一方面對損傷后的細(xì)胞周期有一定影響, 另一方面可能與DNA 損傷修復(fù)偶聯(lián). 有文獻(xiàn)報(bào)道, gadd153對轉(zhuǎn)錄因子C/EBP (CAAT增強(qiáng)子結(jié)合蛋白) 家族有負(fù)調(diào)控作用[10]. 實(shí)驗(yàn)結(jié)果顯示, 經(jīng)紫外線損傷后無論年輕及衰老細(xì)胞出現(xiàn)的G1 期的阻滯同時(shí)伴隨著gadd153、p 21、p 53 基因的高表達(dá), 這說明這三種基因的可誘導(dǎo)與細(xì)胞周期阻滯有著直接的關(guān)系; 然而, 有趣的是衰老細(xì)胞在紫外線照射前即存在明顯的G1 期阻滯, 此時(shí)僅見p 21 的高表達(dá), 這說明p 21對衰老細(xì)胞G1 期的阻滯可能起主要作用, 而p21的高表達(dá)是因?yàn)樗ダ霞?xì)胞中存在的損傷累積不能被有效修復(fù)所致. 因?yàn)樗ダ霞?xì)胞本身即存在明顯的G1期阻滯, 加之p 53、gadd153, 尤其是p 21 基因的誘導(dǎo)性下降, 可能是衰老細(xì)胞G1 期增幅低于年輕細(xì)胞的原因. 同時(shí), 衰老細(xì)胞修復(fù)能力的下降也可能與gadd 基因(包括gadd153, gadd45 等)、p 21 及p 53 等基因的DNA 損傷后的可誘導(dǎo)性在衰老細(xì)胞中的下降有關(guān). 實(shí)驗(yàn)結(jié)果顯示, 衰老細(xì)胞中雖然p 21 這一細(xì)胞周期抑制因子的高表達(dá)導(dǎo)致了細(xì)胞的G1 期阻滯, 為修復(fù)損傷爭取了時(shí)間, 但其它與修復(fù)直接相關(guān)的基因卻不能有效地表達(dá)和發(fā)生作用. 實(shí)驗(yàn)中還發(fā)現(xiàn), p 53 表達(dá)變化并不十分顯著, 說明p 21 在衰老細(xì)胞中的高表達(dá)有可能通過非p 53 途徑調(diào)控[ 10 ]; 但另一方面這可能與p 53 作用通過磷酸化狀態(tài)的改變而實(shí)現(xiàn)有關(guān)[ 11 ] , 在損傷誘導(dǎo)后, p 53 蛋白與各種基因的結(jié)合活性可能通過其磷酸化狀態(tài)的改變而有所改變, 這同樣可能是衰老細(xì)胞中p 53 未見顯著高表達(dá)的原因之一. 然而, 這幾種基因在衰老細(xì)胞中的損傷可誘導(dǎo)性下降卻具有一致性, 這可能正是衰老細(xì)胞在DNA 損傷后細(xì)胞周期調(diào)控能力及修復(fù)能力下降的根本原因之一. 總之, 探討衰老細(xì)胞在DNA 損傷后細(xì)胞周期調(diào)控、DNA損傷修復(fù)以及DNA 損傷可誘導(dǎo)基因的表達(dá)改變, 有助于DNA 損傷修復(fù)基因調(diào)控網(wǎng)絡(luò)的闡明, 對于揭示DNA 損傷修復(fù)與細(xì)胞衰老的關(guān)系具有重要的理論意義.
References
1 童坦君, 張宗玉. 醫(yī)學(xué)老年學(xué)——衰老與長壽. 北京: 人民衛(wèi)生出版社, (Tong Tanjun, Zhang Zongyu. M ed ical Gerontology ——Senescence and long ev ity. Beijing: People’s Health P ress) 1995:139~ 142
2 Paulovich A G, Toczysk iD P, Hartwell L H. When checkpoints fail. Cell, 1997, 88: 315~321
3 Scanchez Y, Elledge S J. Stopped fo r repairs. B ioE ssay s, 1995, 17(6) : 545~ 548
4 南新升, 張宗玉, 黃莉, 張昌潁. 衰老過程中大鼠脾細(xì)胞DNA 修復(fù)能力的變化, 中華老年醫(yī)學(xué)雜志, (N an Xinsheng, ZhangZongyu,Huang L i, Zhang Changying. Change of DNA repair ca2pacity of rat sp leen cells during aging. Ch in J Geriatrics) 1992,11 (5): 301
5 T illey R,M iller S, Srivastava V , Busbee D. Enhanced unsched2uled DNA synthesis by secondary cultures of lung cells estab2lished from calo rically restricted aged rats. Mech Ageing D ev ,1992, 63: 165~ 176
6 王文恭, 童坦君. 細(xì)胞凋亡過程中c2erbB22 基因的表達(dá). 中國生物化學(xué)與分子生物學(xué)報(bào)(W angW engong, Tong Tanjun. Theex2pression of c2erbB22 on apoptotic process. Ch in J B iochem M olB iol) , 1998, 14 (3) : 314~ 317
7 王文恭, 童垣君. 小鼠成纖維細(xì)胞凋亡過程中p53 與bcl22 表達(dá)的時(shí)序性. 中國生物化學(xué)與分子生物學(xué)報(bào)(W ang W engong,Tong Tanjun. The time o rder of p53 and bcl22 gene exp ressionin the apop to tic p rocess of mouse fibroblast cell lines. Ch in JB iochem
M ol B iol) , 1998, 14 (3) : 318~ 321
8 Vo jta P J , Barrett J C. Genetic analysis of cellular senescence.B ioch im B iop hy c A cta, 1995, 1242: 29~ 41
9 Sancar A. DNA repair in humans. A nnu R ev Genetics, 1995, 29:69~105
10 Delmastro D A , L i J , V aisman A , So lle M , Chaney S G. DNAdamage inducible gene exp ression fo llow ing p latinum treatmentin human ovarian carcinoma cell lines. Cancer Chem other P har2m acol, 1997, 39: 245~253
11 V aziri H,W estM D,A llwopp R C, Davison T S,W u Y S, A r2row smith C H, Poirier G G, Bench imo l S. A TM 2dependent telomere loss in aging human dip lo id fibroblasts and DNA dam2age lead to the po st2translational activation of p53 protein in2vo lving po ly (ADP2ribose) polymerase. EM BO J , 1997, 16 (19) :6018~6033




Copyright(C) 1998-2024 生物器材網(wǎng) 電話:021-64166852;13621656896 E-mail:info@bio-equip.com