可靠的CCCadvanced FN1無異源耗材支持人間充質(zhì)干細胞擴增表達
Reliable and Robust Animal-Component-Free hMSC-BM Expansion on Ready-to-Use Eppendorf CCCadvanced™ FN1 Motifs Surface
Aurélie Tacheny¹, Wiâme Ben El Mostapha¹, Nadine Mellies², Françoise De Longueville¹
¹ Eppendorf Application Technologies S.A., Namur, Belgium
² Eppendorf AG, Hamburg, Germany
Abstract
In the last decade, human mesenchymal stem cells (hMSCs) have generated increasing interest in the scientifc world. Many fetal and adult tissues harbor potential multipotent MSCs, which hold a long-term in vitro culturing capacity across several passages without losing their essential characteristics. Prior to their use as a powerful tool for research applications, hMSCs must be expanded in order to reach an adequate number of cells without losing their homing ability, multi-lineage potential, secretion of anti-inflammatory molecules and immunoregulatory effects. Experiments require stable and completely de fned hMSC culture systems consisting of growth surface and culture medium.
The novel Eppendorf CCCadvanced™ FN1 motifs surface represents a completely synthetic cell adhesion-pro moting growth surface for the long-term cultivation of hMSCs in xeno-free and restrictive culture conditions, providing a defned culture system without any animal and human components. This ready-to-use surface is made up of fbronectin-derived motifs to support cell attachment in various serum-free media by mimicking native extra-cellular matrix proteins without additional preparation of the surface.

With its unique properties, the FN1 motifs surface combines convenience with reliable hMSC cultivation: the ready-to-use consumable signifcantly reduces labor time and effort for scientists while offering a fully synthetic growth surface with a high level of consistency during long-term hMSC expansion.
Introduction
First described in the 1970s by Friedenstein as bone marrow-derived fbroblast-like precursors, hMSCs, also known as human mesenchymal stromal cells, consist of a heterogeneous population of multipotent cells which can be easily isolated from various tissues, such as adult bone marrow, adult adipose tissue, dental pulp, fetal and neonatal tissues [1-4]. hMSC isolation and identifcation rely exclu sively on in vitro expanded cell properties. According to the criteria defned by the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy, hMSCs must present ex-vivo plastic-adherent growth abili ties under standard culture conditions and must express a specifc set of cell surface antigens such as CD73, CD90 and CD105, while lacking expression of CD11b, CD19, CD34, CD45 and HLA-DR [5]. Moreover, to conform to minimal hMSC defnition criteria, cells must also be able to differen tiate in vitro into osteogenic, chondrogenic and adipogenic lineages [6].
Besides their characteristic mesodermal differ- entiation potential, it has also been reported that these cells are able to transdifferentiate into additional non-mesodermal cell types including hepatocytes, cardiomyocytes, neuron-like cells and pancreatic-like cells [7]. Very interestingly, these cells present low immunogenicity and possess the ability to secrete soluble bioactive factors that can modulate the im mune system and inflammation process and promote tissue repair [8, 9]. This unique combination of properties makes hMSCs a promising stem cell population in various cell ap plications.
hMSC culture conditions
Present at relatively low abundance in their tissue of ori gin, hMSCs require a robust in vitro cell culture expansion process in order to reach sufcient numbers of high-quality cells. Traditionally, hMSC in vitro expansion occurs in a serum-containing culture system. In the presence of serum associated proteins, hMSC culture can be performed on tis sue culture (TC) treated plastic vessels without any specifc coating.
Nevertheless, the common use of animal-derived materi als, such as serum, represents a non-defned composition, accompanied by variable lot-to-lot quality and purity as well as potential contamination risk, which could be problematic in a wide range of basic and applied research applications [10, 11]. For the past several years, a growing section of the academic and industrial scientifc community has been leaning increasingly towards the use of well-defned, serum free, xeno-free (XF) or animal-component-free (ACF) hMSC culture systems.
More consistent and defned culture conditions are also of great interest to those working with hMSC cultures in con junction with biopharmaceutical production, drug screen ing or disease modeling, as these felds require robust cell performances with a high level of consistency and repro ducibility. In the absence of serum proteins, hMSCs expand ed in serum-free culture systems necessitate additional cell adhesion-promoting coating on the culture surface. To ensure the defned nature of the culture system, coatings of biologi cal origin should be excluded.
Synthetic FN1 motifs surface
Based on a proprietary coating technology, the Eppendorf CCCadvanced FN1 motifs surface consists of synthetic fbronectin-derived motifs (including RGD), specifcally designed to mimic the cell attachment site of native extra cellular matrix (ECM) proteins. Used in combination with synthetic culture medium and dissociation solution, this surface represents an effective animal- and human-compo nent–free alternative to other biological coating-dependent culture systems.
Being ready-to-use, it constitutes a real improvement for stem cell researchers, signifcantly reducing labor time and effort while offering lot-to-lot consistency and more reliable performance in comparison with self-coating solutions. The FN1 motifs surface is suitable for the long-term expansion of bone marrow-derived hMSCs (hMSC-BM), and it is com patible with various commercial xeno-free media.
Cultured on this novel synthetic surface, hMSC-BM main tain their characteristic cell morphology, as well as linear cumulative population doubling, without the appearance of replicative senescence-associated signs, across 10 succes sive passages. Following successive passages in a completely defned animal-component-free culture system, cells which were expanded on the FN1 motifs surface maintain a typical hMSC surface antigen marker expression profle and the ability to differentiate in vitro into osteoblasts, adipocytes and chondrocytes, representing the three mesoderm lineages.
Materials and Methods
Short-term cell growth evaluation
Lonza™ Poietics™ human mesenchymal stem cells (hMSC-BM, PT-2501, Lonza) derived from normal adult bone marrow were thawed at passage 2 and pre-cultivated as recommended by the supplier. In order to evaluate the ability of the Eppendorf CCCadvanced FN1 motifs surface to efciently support hMSC-BM short-term expansion under different xeno-free (XF) culture conditions, hMSC-BM (P3) were plated at the initial cell density of 3,500 cells/cm2, either on a TC treated culture surface or on the ready-to-use FN1 motifs surface in the presence of 3 different XF culture media: StemPro® MSC SFM XenoFree medium (A1067501, ThermoFisher Scientifc®), MesenCult™ ACF Medium (05449, STEMCELL™ Technologies) and Corning® stemgro® hMSC Medium (40-410-KIT, Corning). In parallel, cells were similarly plated on both surfaces in a serum-containing medium (MSCGM™, PT-3001, Lonza). After seeding in the appropriate culture medium, hMSCs were incubated under standard cell culture conditions (37 °C, 5% CO2, humidifed atmosphere) and fed every 3 days by refreshment of the entire volume of culture medium. At day 4 and day 7 post seeding, after microscope analysis, cells were harvested with 0.25% Trypsin/EDTA according to manufacturer’s recommendations. After centrifugation, three independent cell counts were performed on each cell suspension using the Vi-CELL™ automated cell counting device (Beckman Coulter®, USA). The average viable cell density was then used to evaluate the viable cell number/cm2 in each of the growth conditions.
Long-term cell growth evaluation
A unique pool of human bone marrow-derived mesenchymal stem cells (hMSC-BM, PT-2501, Lonza) was thawed at pas sage 2 and seeded directly at the initial cell density of 3,500 cells/cm2, either on the FN1 motifs surface or on two other synthetic culture surfaces, the ready-to-use surface by Competitor A and the surface by Competitor B which offers a self-coating solution. In order to ensure a com pletely defned animal-component-free (ACF) culture system during the entire expansion process, cells were expanded in StemPro MSC SFM XenoFree medium and harvested for subculture using TrypLE™ Select 10x (A12177-01, Thermo Fisher Scientifc). As a reference, hMSC-BM were expanded in parallel in a traditional culture system consisting of a TC treated surface, 10% FBS-containing culture medium and 0.25% Trypsin/EDTA as detachment solution. After seeding in the appropriate culture medium, hMSC-BM were incubat ed under standard cell culture conditions (37 °C, 5% CO2, humidifed atmosphere) and fed every 3 days by refresh ment of the entire culture medium volume until a confluence level of 70-80% was reached.
For each experimental condition, cells were cultured for 10 successive passages (from P3 to P12). At each passage, cell morphology was examined with the EVOS® FL Cell Im aging System (Thermo Fisher Scientifc, USA). Cell growth and viability were assessed on 3 independent T75 flasks per experimental condition. After complete cell detach ment, a cell count was performed on each homogenized cell suspension using the Vi-CELL automated cell counting device (Beckman Coulter, USA). Population doubling (PD) and doubling time (DT) were calculated using the respective formula:
PD = (log10(NH)-log10(Ni))/log10(2)
DT = time in culture (hours) x (LN(2)/ LN(NH/Ni))
NH = total number of harvested viable cells
Ni = initial number of seeded cells
For the purpose of evaluating statistical significance, a Student t-test was performed on normalized doubling time data ob tained during 10 successive passages.
β-galactosidase staining
At every two passages within the expansion process, the senescence-associated β-galactosidase (SA-β-gal) activity was evaluated on expanded hMSC-BM using the Senes cence Cells Histochemical Staining Kit (CS0030, Sigma) in accordance with the manufacturer’s instructions.
hMSC-BM surface marker expression analysis by flow cytometry
The preservation of the hMSC-specifc immunophenotype was assessed on the initial cell population (input cells) and at the end of the long-term expansion process for each experimental condition. The positive and negative expression levels of several key surface markers (positive markers: CD44, CD73, CD90 and CD105; negative markers: CD11b, CD19, CD34, CD45, and HLA-DR) were evaluated through flow cytometry analyses using the BD Stemflow™ Human MSC Analysis Kit (562245, BD Biosciences). Briefly, viable cell density was determined from a single-cell suspension via cell count, and the appropriate number of cells (input cells: 10,000) was prepared for FACS analyses according to the procedure recommended by the antibody kit manufacturer. For each cell type analyzed, a sample of unstained cells, as well as an isotype control, were prepared in order to measure auto-fluorescence and non-specifc staining, respectively. Cells were analyzed with a BD FACSVerse flow cytometer (BD Biosciences, USA), and data analysis was performed using the BD FACSUITE™ SOFTWARE (BD Biosciences, USA).
hMSC-BM multi-lineage differentiation potential
Preservation of the multipotent differentiation potential was evaluated on hMSC-BM expanded for 5 passages on the FN1 motifs surface in an ACF culture system. Osteo genic, adipogenic and chondrogenic differentiation was induced by using the MesenCult™ Osteogenic Stimulatory Kit (Human) (05434, STEMCELL Technologies), the hMSC Adipogenic Differentiation Bulletkit™ (PT-3004, Lonza) or the hMSC Chondrogenic Differentiation Bulletkit (PT-3003, Lonza), respectively, according to the manu facturer’s instructions. As negative controls, uninduced cells were maintained in parallel in their initial culture medium. Respective differentiation efciencies were assessed by specifc stainings. The osteogenic differentiation and mineralized matrix accumulation were highlighted by Alizarin Red staining performed 21 days post-induction. The intracellular lipid droplet accumulation associated to the adipogenic differentiation was confrmed by Oil Red O staining per formed 21 days post-induction and the glycosaminoglycans secreted by chondrocytes were observed through Alcian blue specifc staining performed 14 days post-induction.
Results and Discussion
The Eppendorf CCCadvanced™ FN1 motifs surface supports an efcient short-term expansion of hMSC-BM in various xeno-free culture media The ready-to-use FN1 motifs surface supports efcient hMSC-BM growth in combination with different serum-free and xeno-free culture media.
The morphology and proliferation of hMSCs were evaluated 7 days post-seeding on the FN1 motifs surface and on a TC treated surface, in different commercial culture media, respectively (Figure 1 and 2). In traditional serum-contain ing culture systems, hMSC-BM adhered and pro-liferated similarly on the FN1 motifs and the TC treated surface. On both surfaces, cells exhibited their typical fbroblast-like morphology after short-term expansion. In the absence of serum in the culture medium, hMSCs had difculty adher ing and proliferating on the TC treated surface, suggesting the need for additional cell adhesion-promoting coating. By contrast, regardless which of the XF media were tested, the FN1 motifs surface efciently supported hMSC-BM attachment and growth.
As previously described in the literature and by media sup pliers, cells expanded under xeno-free culture conditions exhibited more elongated spindle-shaped cell morphology as compared to cells expanded in the presence of serum and were associated with more compact mono-layers and higher cell densities [12, 13]. Starting from a comparable initial cell density, hMSC-BM yields obtained 7 days post-seeding on the FN1 motifs surface in the three tested XF culture media indicate efcient and similar cell expansion. These results confrm the suitability of the FN1 motifs surface for xeno free expansion of hMSC-BM in combination with various commercial media.
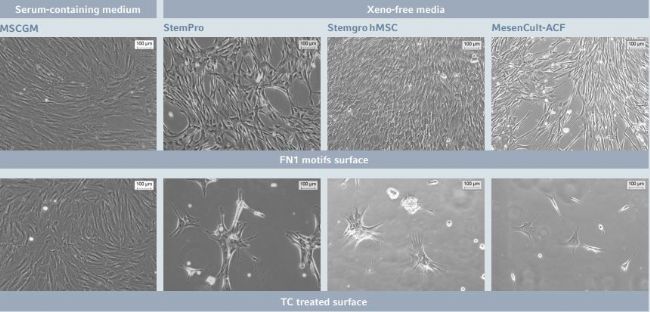
Figure 1: hMSC-BM morphology after short-term expansion on Eppendorf CCCadvanced FN1 motifs surface in different culture media
hMSCs cultured in traditional serum-containing medium display their characteristic fbroblast-like morphology after short-term expansion on both the FN1 motifs surface and a TC treated surface. In contrast to the TC treated surface, the FN1 motifs surface also efciently supports hMSC-BM attachment and growth in various serum-free culture systems, showing a more elongated spindleshaped cell morphology, more compact monolayers and higher cell densities. The images show representative areas at 7 days post-seeding. Scale bar indicates 100 µm.
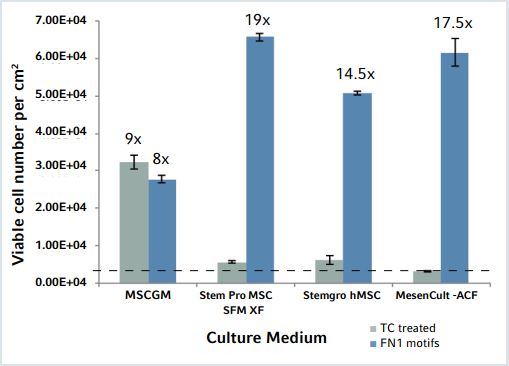
Figure 2: hMSC-BM proliferation after short-term expansion on Eppendorf CCCadvanced FN1 motifs surface in different culture media
Seeded at comparable cell densities in different commercial serumfree media, the FN1 motifs surface efciently supports xeno-free hMSC expansion with high viable cell numbers 7 days post-seeding. Results are expressed in viable cell number per cm2 and represent a mean of three independent cell counts performed 7 days post-seeding. The dotted line indicates the initial cell seeding density (3,500 cells/ cm2). Average fold inductions are noted above columns.
The Eppendorf CCCadvanced™ FN1 motifs surface supports an efcient long-term expansion of hMSC-BM in a completely defned, animal-component-free culture system
hMSCs were maintained for 10 successive passages on the FN1 motifs surface in order to confrm that this synthetic culture surface supports long-term hMSC-BM expansion in a completely defned, animal-component-free culture system without impacting cell quality. In parallel, hMSC-BM were maintained in an identical ACF culture system on two competitor surfaces, the ready-to-use surface by Competitor A and the self-coated surface by Competitor B. Moreover, cells expanded in a traditional culture system (TC treated surface, serum-containing culture medium and Trypsin/ EDTA as detachment solution) were used as a reference. The morphology of the hMSC-BM was monitored at each passage, under each experimental condition, during the entire expansion process of 10 successive passages, and it was recorded at P4 and P12, respectively, after 2 and 10 successive passages on the different cell culture surfaces (Figure 3).

Figure 3: hMSC-BM morphology after long-term expansion on Eppendorf CCCadvanced FN1 motifs surface and competitor surfaces in animal-component-free conditions
hMSC-BM were expanded for 10 successive passages on the FN1 motifs surface, a ready-to-use surface by Competitor A and a self-coated surface by Competitor B, each showing elongated cells characteristic of hMSC expansion under ACF culture conditions. However, long-term expansion on the surface by Competitor A promoted the formation of large hMSCs such as those observed in a traditional culture system (TC treated surface – serum-containing medium – Trypsin/EDTA), whereas the FN1 motifs surface preserve the characteristic hMSC morphology in restrictive culture conditions during the successive 10 passages. The images show representative areas at the respective passages. Scale bar indicates 400 µm.
At an early passage following seeding (P4), cells displayed the expected hMSC-BM morphology corresponding to a fbroblast-like, spindle-shaped morphology on each tested synthetic surface. As previously described, cells expanded in a serum-containing culture system presented a wider spin dle-shape as compared to the more elongated cells expanded in serum-free conditions. On the FN1 motifs surface, this mor phology remained stable across passages. By contrast, cells expanded in the traditional culture system or on the competi tor A surface exhibited a higher proportion of very large cells at P12, suggesting the emergence of replicative senescence signs [14].
The altered cell morphology was indeed associated with a progressive increase of senescence-associated β-galactosidase positive cells as well as with a decreased proliferation potential during progressive hMSC expansion on a TC treated surface in the serum-containing culture system, especially after passage 8 (Figure 4). By contrast, hMSCs expanded in ACF culture conditions on the FN1 motifs surface maintained a stable doubling time across passages with no signifcant β-galactosidase activity.
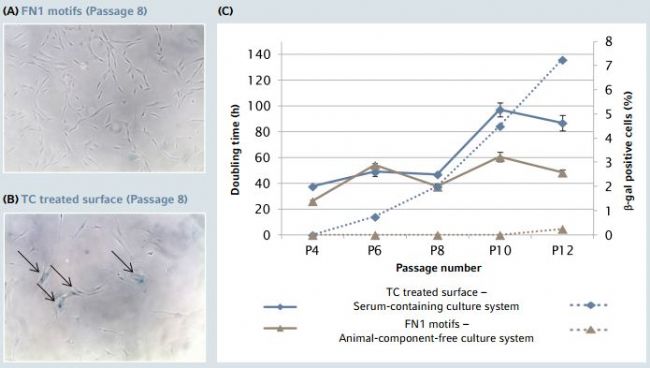
Figure 4: hMSC replicative senescence characterization during long-term expansion on Eppendorf CCCadvanced FN1 motifs surface
(A/B) While β-galactosidase staining revealed an increasing number of positive hMSCs in a traditional culture system (TC treated surface – serum-containing medium – Trypsin/EDTA), indicating a permanent cell-cycle arrest from passage 8, hMSCs expanded on the FN1 motifs surface in animal-component-free medium exhibited no senescent phenotype at the same stage. The images show representative areas after β-galactosidase staining of hMSCs, and arrows indicate β-galactosidase positive cells. (C) Expansion on the FN1 motifs surface prevented senescence as characterized by the absence of signifcant β-galactosidase activity and a stable doubling time across passages. Solid lines illustrate doubling times calculated from the proliferation assay. Dotted lines illustrate the percentage of β-galactosidase positive cells. Results represent a mean of three independent cell counts (n=3).
The FN1 motifs surface supported robust and stable hMSC proliferation during the entire culture period of 10 successive passages (Figure 5). Compared with results obtained on other growth surfaces used in combination with animal component-free conditions, the culture of hMSC-BM expand ed on FN1 motifs surface achieved a signifcantly faster pro liferation rate, as shown by a short doubling time and high population doubling number. These results demonstrate that the FN1 motifs surface is a ready-to-use synthetic surface for efcient hMSC expansion in ACF culture conditions.
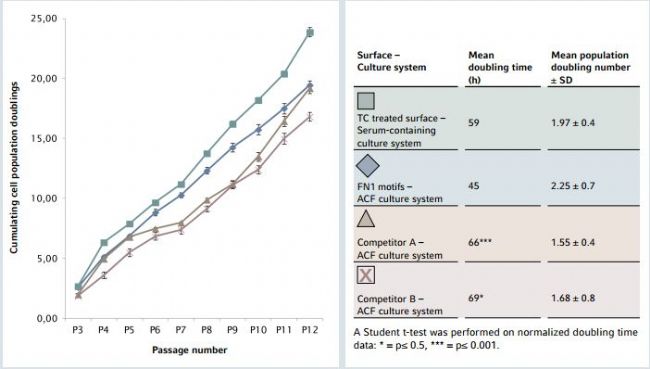
Figure 5: hMSC proliferation rate during long-term expansion in different animal-component-free culture systems
hMSC expansion on the FN1 motifs surface ensured a signifcantly faster proliferation rate with a short doubling time and high population doubling number in animal-component-free conditions. Results are expressed as cumulative cell population doubling numbers over 10 successive passages on the FN1 motifs surface and two competitor surfaces in an ACF culture system. Cells expanded on a TC treated surface in a traditional serum-containing culture system served as a reference. Results represent a mean of three independent cell counts (n=3). For each experimental condition, the mean cell population doubling time and the mean population doubling number per passage are indicated.
Even after long-term expansion on the FN1 motifs surface, hMSC-BM maintained their specifc immunophenotype and continued to express high levels of mesenchymal markers such as CD105, CD73, CD90 and CD44, while lacking the expression of hematopoietic lineage markers (< 1% CD11b, CD34, CD45, CD79 α, and HLA-DR positive hMSCs in the entire cell population) according to the standard criteria for MSC identifcation recommended by the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy (Figure 6) [5]. The hMSC marker expression profle obtained on the FN1 motifs surface is comparable to other animal-component-free culture systems as well as a traditional serum-containing culture system (Table 1).
Table 1: Comparison of hMSC-specifc cell surface antigen expression profle after long-term expansion in different culture systems.
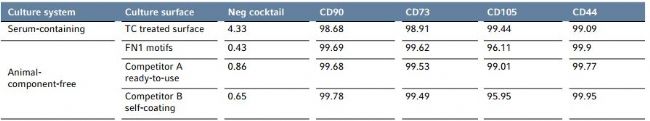
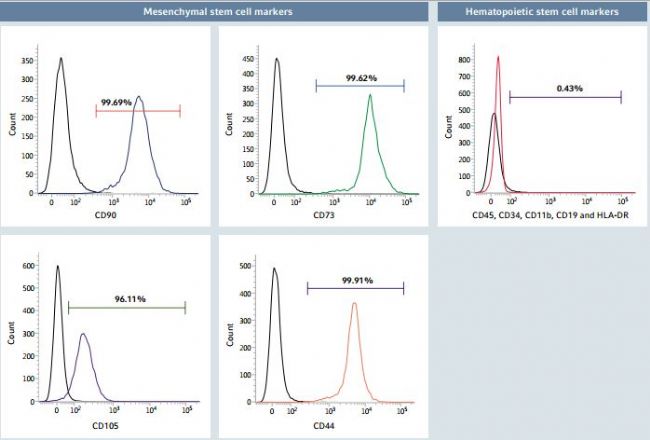
Figure 6: Flow cytometry analysis of hMSC markers after long-term expansion on Eppendorf CCCadvanced FN1 motifs surface in an animal-component-free environment
After long-term expansion across 10 successive passages on the FN1 motifs surface in animal-component-free medium, hMSCs showed a typical expression profle of characteristic surface markers as evaluated by flow cytometry analysis: more than 95% of cells of the total cell population expressed the mesenchymal markers CD90, CD73, CD105 and CD44, while lacking the expression of hematopoietic surface markers CD45, CD34, CD11b, CD19 and HLA-DR. Unstained cells and isotype controls were prepared in order to validate staining specifcity. Isotype controls (in black) were used to determine the percentage of positive cells for each of the markers of interest.
The functional multipotency of hMSCs after long-term ex pansion on the FN1 motifs surface in an ACF culture system was ultimately confrmed by their in vitro differentiation into cells of three mesoderm lineages, the osteogenic, adipogenic and chondrogenic lineages (Figure 7).
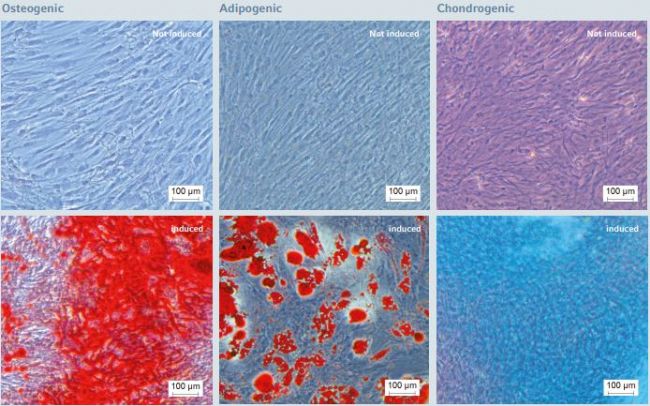
Figure 7: Multi-lineage differentiation potential of hMSC-BM after long-term expansion on the Eppendorf CCCadvanced FN1 motifs surface in an animal-component-free environment
After 5 successive passages on the FN1 motifs surface under animal-component-free conditions, hMSCs maintained their multi-lineage differentiation potential as examined by specifc fluorescent staining of three specifc mesoderm lineages, respectively: hMSCs successfully differentiated into osteogenic (Alizarin Red staining – 21 days post-induction), adipogenic (Oil Red O staining – 21 days post-induction) and chondrogenic (Alcian blue staining – 14 days post-induction) lineages. Scale bar indicates 100 µm.
Conclusion
The ready-to-use Eppendorf CCCadvanced FN1 motifs sur face efciently supports long-term hMSC-BM expansion in a completely defned, animal-component-free culture system. During the expansion process across 10 successive passages hMSCs maintain a stable and robust proliferation rate with their characteristic morphology without signs of replicative senescence. The undifferentiated hMSCs retain their typical marker expression profle as well as their multi-lineage mesodermal differentiation potential. The suitability of the FN1 motifs surface to support efcient hMSC-BM proliferation in different commercial xeno-free culture media facilitates the establishment of an animal-component-free environment for hMSC cultivation.
Literature
[1] Friedenstein ZJ, Deriglasova UF, Kulagina NN, Panasuk AF, Rudakowa SF, Luriá EA, Ruadkow IA. Precursors for fbroblasts in different populations of hematopoietic cells as detected by the in vitro colony assay method. Experimental Hematology 1974; 2(2): 83-92.
[2] Horwitz EM, Le Blanc K, Dominici M, Mueller I, Slaper-Cortenbach I, Marini FC, Deans RJ, Krause DS, Keating A; International Society for Cellular Therapy. Clarifcation of the nomenclature of MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005; 7(5): 393-395.
[3] Bianco P, Riminucci M, Gronthos S, Robey PG. Bone marrow stromal stem cells: Nature, biology and potential applications. Stem Cells 2001; 19(3): 180–192.
[4] Bobis S, Jarocha D, Majka M. Mesenchymal stem cells: characteristics and clinical applications. Folia Histochemica et Cytobiologica 2006; 44 (4): 215-230.
[5] Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop Dj, Horwitz E. Minimal criteria for defning multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006; 8(4): 315–317.
[6] Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science 1999; 284(5411): 143–147.
[7] Phinney DG and Prockop DJ. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair – Current views. Stem Cells 2007; 25(11): 2896-2902.
[8] Bernado ME, Pagliara D, Locatelli F. Mesenchymal stromal cell therapy: a revolution in regenerative medicine? Bone Marrow Transplantation 2012; 47(2): 164-171.
[9] Wang Y, Chen X, Cao W , Shi Y. Plasticity of mesenchymal stem cells in immunomodulation: pathological and therapeutic implications. Nature Immunology 2014; 15: 1009–1016.
[10] Kinzebach S and Bieback K. Expansion of mesenchymal stem/stromal cells under xenogenic-free culture conditions. Advances in Biochemical Engineering/ Biotechnology 2013; 129: 33-57.
[11] Karnieli O, Friedner OM, Allickson JG, Zhang N, Jung S, Fiorentini D, Abraham E, Eaker SS, Yong TK, Chan A, Grifths S, When AK, Oh S, Karnieli Ol. A consensus introduction to serum replacements and serum-free media for cellular therapies. Cytotherapy 2017; 19(2): 155-169.
[12] Li CY. Wu XY, Tong JB, Yang XX, Zhao JL, Zheng QF, Zhao GB, Ma ZJ. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Research and Therapy 2015; 6:55.
[13] Dolley-Sonneville PJ, Romeo LE, Melkoumian ZK. Synthetic surface for expansion of human mesenchymal stem cells in xeno-free, chemically defned culture conditions. PLoS One 2013; 8(8):e70263.
[14] Wagner W, Horn P, Castoldi M, Diehlmann A, Bork S, Saffrich R, Benes V, Blake J, Pfster S, Eckstein V, Ho AD. Replicative senescence of mesenchymal stem cells: a continuous and organized process. PLoS One 2008; 3(5):e2213.