
代謝組學(xué)助力探究3-苯乳酸對引起腐爛百合鱗莖的米根霉菌的抑制作用
文章標(biāo)題:Energy metabolism as the target of3-phenyllactic acid against Rhizopus oryzae
發(fā)表期刊:InternationalJournal of Food Microbiology
發(fā)表期刊:InternationalJournal of Food Microbiology
影響因子:5.277
合作客戶:蘭州理工大學(xué)
百趣生物提供服務(wù):TMT+中心碳
分組:2mg/mLPLA治療組、4mg/mLPLA治療組、8mg/mLPLA治療組和對照組(n=3)
研究背景
3-苯乳酸(PLA)具有廣泛的抗真菌活性,但對PLA在真菌細(xì)胞上的作用靶點及其作用機制的研究較少。本研究探討了3-苯乳酸對引起腐爛百合鱗莖的米根霉菌(R.oryzae)的抑制作用。PLA對米根霉菌的最低抑菌濃度為8mg/mL。通過掃描電鏡和透射電鏡觀察米根霉菌的超微結(jié)構(gòu),結(jié)果表明PLA對米根霉菌的細(xì)胞膜沒有損傷,但對線粒體和其他細(xì)胞器有破壞作用。TMT蛋白組分析結(jié)果顯示PLA顯著下調(diào)己糖激酶(HK)、磷酸果糖激酶(PFK)、a-酮戊二酸脫氫酶(a-KGDH)、腺苷酸激酶(ADK1)、細(xì)胞色素C氧化酶(Cytochrome C oxidase)和NADH脫氫酶(NADH dehydrogenase)的表達(dá)(P<0.05)。在糖酵解、三羧酸循環(huán)或氧化磷酸化代謝過程中,線粒體ADP/ATP載體蛋白(AAC)和亞基IV(CCIO IV)表達(dá)上調(diào)(P<0.05)。在此基礎(chǔ)上,通過酶活性檢測發(fā)現(xiàn)上述通路HK和a-KGDH活性下調(diào),并通過實時熒光定量PCR進(jìn)一步證實ADK1、AAC、CCIO IV和NADH脫氫酶基因表達(dá)下調(diào)。中心碳代謝組學(xué)研究表明,三羧酸循環(huán)代謝產(chǎn)物中檸檬酸、順-烏頭酸、異檸檬酸、-酮戊二酸、琥珀酸、富馬酸和蘋果酸均顯著下調(diào)(P<0.05),氧化磷酸化產(chǎn)生的ATP也顯著減少(P=0.02)。導(dǎo)致能源生產(chǎn)不足。因此,ROS水平升高了141%,細(xì)胞色素C被釋放,細(xì)胞逐漸凋亡。結(jié)果表明,能量代謝是PLA對米根霉的作用靶點。這一研究首次表明PLA對米根霉的能量代謝可能是其作用靶點,為進(jìn)一步研究其抑菌機理提供了理論依據(jù)。
百趣代謝組學(xué):機制探索不發(fā)愁,淺看真菌防治靶點代謝組學(xué)研究思路
研究結(jié)果
3-苯乳酸對米根霉生長的抑制作用
PLA處理組(2和4mg/mL)菌體的菌絲干重均低于對照組。當(dāng)PLA濃度達(dá)到8 mg/mL時,隨著培養(yǎng)時間的延長,米根霉測得的干重基本沒有變化,說明在該濃度下米根霉的生長受到了嚴(yán)重的抑制。實驗結(jié)果表明,3-苯乳酸對米根霉的生長有較好的抑制作用,且隨著3-苯乳酸濃度的增加,對米根霉的抑制作用越明顯。
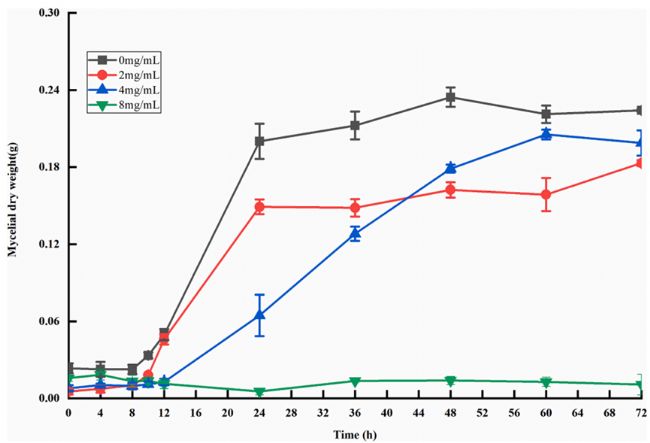
圖1. 米根霉的生長曲線(灰色線為對照組,紅色線為2mg/mLPLA治療組
3-苯乳酸對菌絲形態(tài)和超微結(jié)構(gòu)的影響
用掃描電鏡(SEM)觀察了稻瘟菌的菌絲形態(tài)。對照樣品形態(tài)正常,菌絲飽滿,表面光滑(圖2.A1和A3)。PLA處理米根霉24h后,我們發(fā)現(xiàn)菌絲密集雜亂(圖2.A2和A4),菌絲管腫脹起皺;但表面無明顯損傷或缺失(圖2.A4)。在菌絲表面觀察到一些分泌物(圓圈),推測為胞外多糖(圖2.A4)。因此,推斷PLA處理對米根霉細(xì)胞壁沒有造成嚴(yán)重的破壞。
電鏡觀察顯示,對照組米根霉的超微結(jié)構(gòu)完整,細(xì)胞壁正常光滑,細(xì)胞膜結(jié)構(gòu)正常,胞漿壁無分離,線粒體形成相對規(guī)則(圖2.B1-B4)。經(jīng)3-苯乳酸處理后,整個細(xì)胞結(jié)構(gòu)萎縮,細(xì)胞外壁變薄,厚度不均勻。內(nèi)側(cè)細(xì)胞膜呈齒狀凹陷,局部漿壁分離。細(xì)胞核消失,基質(zhì)和內(nèi)嵴不可見。細(xì)胞膜呈齒狀向內(nèi),漿壁部分分離(圖2.B4中紅色箭頭)。線粒體固縮,液泡塌陷。
綜上所述,PLA對米根霉線粒體及其他細(xì)胞器的破壞程度較高,這可能是PLA對米根霉抑制作用的靶點。
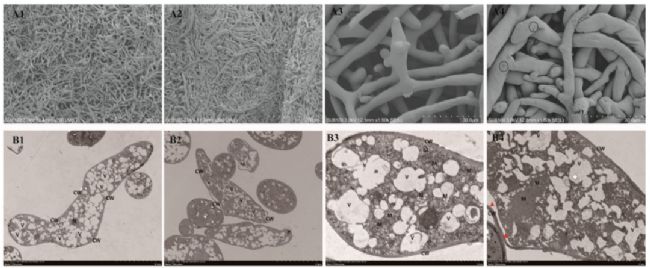
圖2.對照和PLA處理米根霉的SEM和TEM檢查:(A)SEM檢查,(B)TEM檢查
3-苯乳酸的蛋白表達(dá)對米根霉菌有抑制作用
利用TMT蛋白質(zhì)組學(xué)方法,在PLA處理組和對照組之間發(fā)現(xiàn)了485個差異表達(dá)的蛋白。其中,PLA處理組有205個蛋白表達(dá)上調(diào),280個蛋白表達(dá)下調(diào)。對差異表達(dá)蛋白進(jìn)行COG分析(圖3)得知PLA可通過控制能量代謝影響米根霉菌的生長。且經(jīng)PLA處理后的米根霉菌中HK、FK、SDH和a-KGDH的活性受到抑制。相比之下,PLA處理后的米根霉菌中PFK活性略有增加,碳流可能被轉(zhuǎn)移到其他丙酮酸分支點。這些結(jié)果與差異蛋白表達(dá)是一致的。
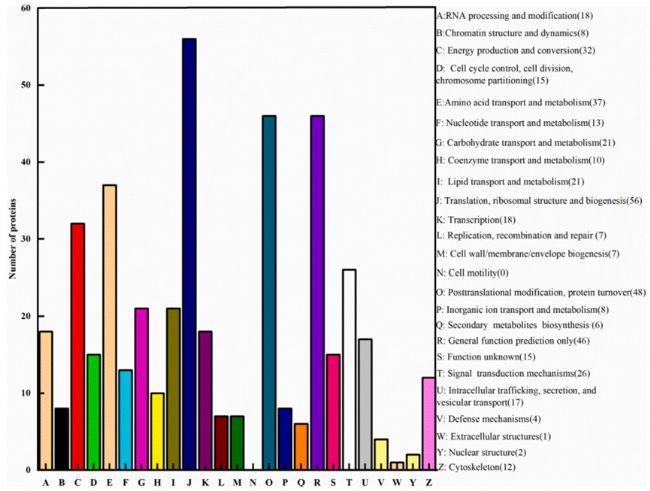
圖3. 差異表達(dá)蛋白的COG分析
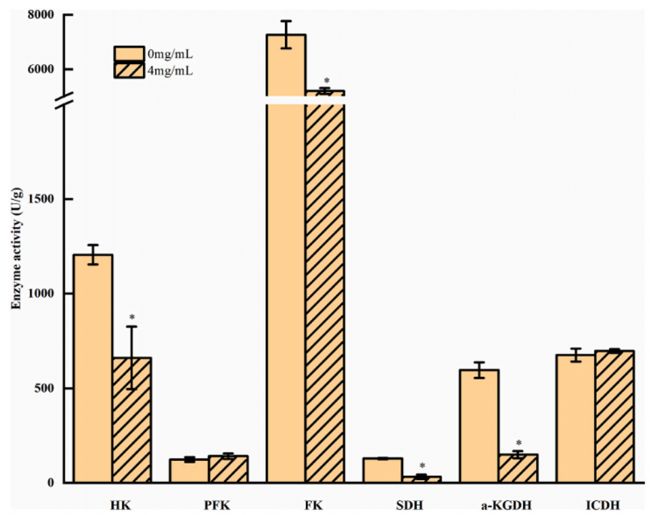
圖4. 與糖酵解和TCA循環(huán)相關(guān)的關(guān)鍵酶活性,星號表示樣本間差異顯
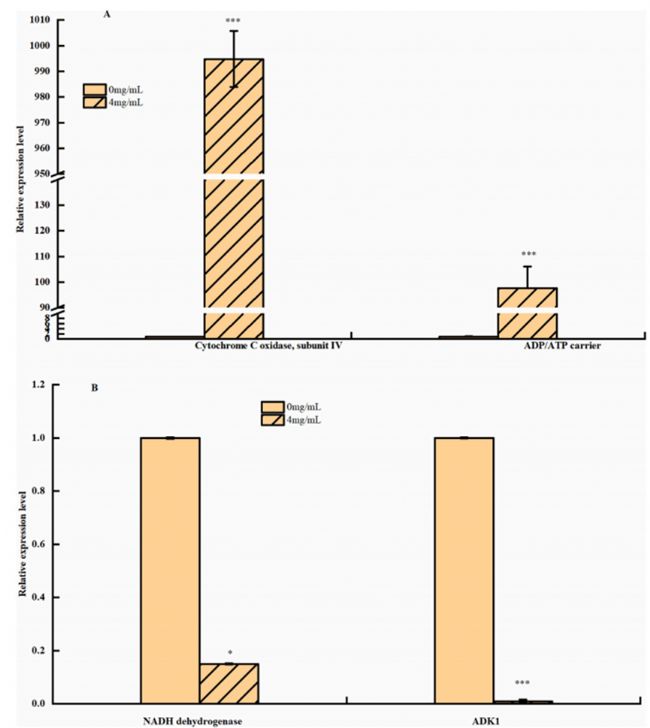
表1. 4mg/mLPLA處理組與對照組在糖酵解、三羧酸循環(huán)或氧化磷酸化
米根霉菌中心碳代謝產(chǎn)物的差異及分析
通過中心碳檢測,我們在對照組和PLA處理組之間確定了51個中心碳代謝產(chǎn)物,其中22個代謝產(chǎn)物顯著上調(diào),29個代謝產(chǎn)物顯著下調(diào) (圖6)。利用KEGG數(shù)據(jù)庫對差異代謝產(chǎn)物的代謝途徑進(jìn)行分析,差異表達(dá)的代謝產(chǎn)物在TCA循環(huán)、乙醛酸和二羧酸代謝、糖酵解或糖異生、丙酮酸代謝、光合生物碳固定和磷酸戊糖途徑中顯著富集。在本研究中, PLA處理顯著降低了米根霉菌中ADP和ATP的水平(表2),說明PLA處理影響了米根霉菌的能量代謝。機體線粒體中的OXPHOS負(fù)責(zé)90%以上的細(xì)胞ATP的產(chǎn)生,結(jié)合蛋白質(zhì)組學(xué)分析,這種效應(yīng)可能是由于OXPHOS中一種或多種酶活性明顯缺乏,導(dǎo)致ATP合成減少。
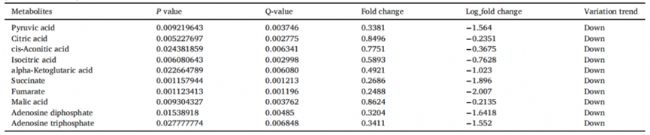
表2. 4mg/mLPLA處理組與對照組在糖酵解、三羧酸循環(huán)或氧化磷酸化
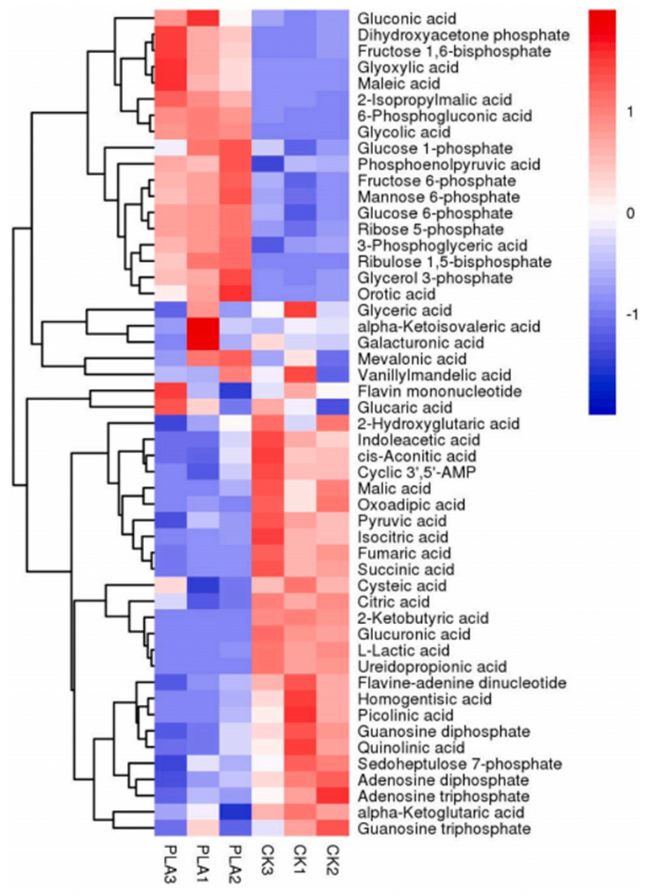
圖6. 中心碳代謝產(chǎn)物在對照組和PLA處理組之間的差異
3-苯乳酸對細(xì)胞色素C釋放和ROS水平的影響
經(jīng)過PLA處理的米根霉菌細(xì)胞質(zhì)中的細(xì)胞色素C含量更高(圖7A),這表明位于線粒體內(nèi)外膜的細(xì)胞色素C缺失。在之前的一項研究中,反式肉桂醛處理導(dǎo)致細(xì)胞色素C滲漏并觸發(fā)線粒體介導(dǎo)的細(xì)胞凋亡。推斷PLA可能導(dǎo)致線粒體呼吸鏈功能障礙,進(jìn)而導(dǎo)致ATP缺乏,導(dǎo)致細(xì)胞死亡。
當(dāng)PLA處理米根霉菌,ROS水平增加,在4.0mg/LPLA時,ROS水平增加了141%(圖7B)。結(jié)果表明,PLA處理誘導(dǎo)水稻植株ROS積累,可能引起線粒體功能障礙和細(xì)胞氧化損傷。
在本研究中,我們發(fā)現(xiàn)PLA影響糖酵解、TCA循環(huán)和氧化磷酸化,導(dǎo)致能量生產(chǎn)不足和細(xì)胞凋亡。雖然目前還沒有關(guān)于對米根霉菌的抑制作用的研究,但與乳酸相比,PLA對其他真菌如黑曲霉具有顯著的抑制作用,與乙酸相比,PLA沒有明顯的刺激性氣味。此外,使用乳酸作為催化劑生產(chǎn)PLA可能是一種可行且環(huán)保的方法。因此,PLA可以作為一種很有前景的天然機制來控制食品中的污染物。
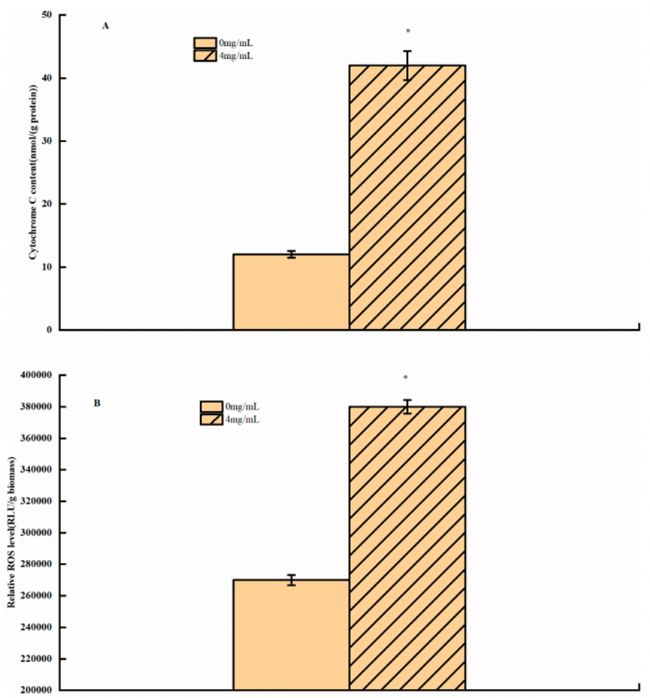
圖7. 對照組和PLA處理組之間的ROS水平和Cyt C釋放,星號表示樣本
結(jié)論
PLA對真菌細(xì)胞靶點的作用及其抗真菌機制研究較少。本研究通過生理生化、電鏡、TMT蛋白質(zhì)組學(xué)和中心碳代謝組學(xué)等方法,探討了米根霉菌對百合腐爛球莖的抑制作用。首先,我們證明了PLA抑制了米根霉菌的生長,破壞了米根霉菌的形態(tài)和超微結(jié)構(gòu)。其次,我們發(fā)現(xiàn)PLA處理降低了與糖酵解和TCA循環(huán)相關(guān)的關(guān)鍵酶的活性,導(dǎo)致丙酮酸和TCA循環(huán)產(chǎn)物的減少和能量的損失。最后,PLA處理通過影響電子傳遞鏈中的關(guān)鍵蛋白來影響氧化磷酸化,導(dǎo)致能量生產(chǎn)不足,且使米根霉菌的ROS水平升高,細(xì)胞色素C釋放,導(dǎo)致細(xì)胞凋亡逐漸發(fā)生。綜上所述,PLA通過能量代謝抑制米根霉菌的生長(圖8)。這是首次對PLA抑制真菌的機制進(jìn)行全面研究。因此,PLA可作為一種有效的天然抗真菌藥物。
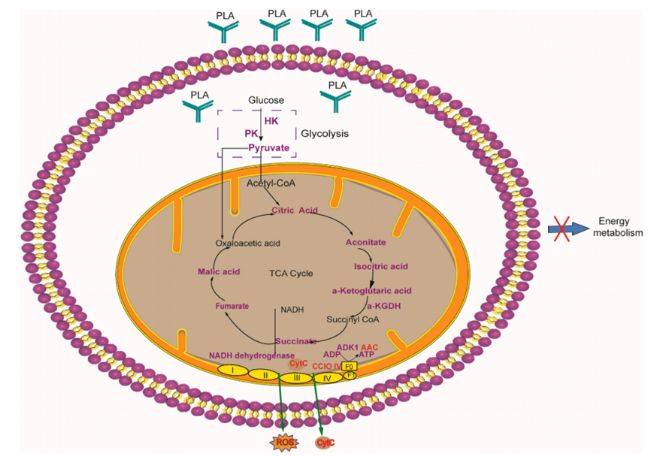
圖8. PLA抑制稻瘟病菌的可能機制是基于能量代謝,紫色代表減少

- MeRIP-seq等揭示m6A甲基化及調(diào)控因子在食管胃結(jié)合部腺癌中的作用
- 單細(xì)胞甲基化多組學(xué)分析揭示哺乳期母體低蛋白飲食對子代的跨代傳遞
- WGBS在揭示主動DNA去甲基化缺失植物的DNA甲基化跨代增加中的應(yīng)用
- SNP分型常見問題及解決方案
- ACE等揭示產(chǎn)前不良環(huán)境暴露通過DNA羥甲基化變化介導(dǎo)子代自閉癥
- 文獻(xiàn)解讀:DNA甲基化保護早期胚胎線粒體基因組穩(wěn)定性
- 血液Target-BS揭示靶向DNA甲基化標(biāo)記水平與患者乳腺癌風(fēng)險相關(guān)
- 多組學(xué)測序分析揭示m5C甲基化上調(diào)E2F1表達(dá)以促進(jìn)卵巢癌腫瘤進(jìn)展



Copyright(C) 1998-2024 生物器材網(wǎng) 電話:021-64166852;13621656896 E-mail:info@bio-equip.com